ORIGINE ET SEPARATION DES ESPECES
Par Robert MAZEL
Revue de l’Association Roussillonnaise d’Entomologie - Supplément 1995.
Préface
Par le Président de
l’Association
Définir l’espèce est un défit pour le naturaliste. Les tentatives se sont succédées et nos illustres prédécesseurs ont proposé des définitions qui ne manquent ni d’audace ni de finesse d’analyse.
Linné était bref : « il y a autant d’espèces qu’en créa à
l’origine l’Etre Infini » !...
Mais les anciens avaient fondé la notion d’espèce avant
tout sur la ressemblance des enfants avec les parents; Cuvier avait repris cette idée et proposait : « la collection de tous les êtres organisés descendus l’un de l’autre ou de parents communs et de
ceux qui leur ressemblent autant qu’ils se ressemblent entre eux ».
Cette définition supposait
la fixité de l’espèce, c’est à dire l’invariabilité des caractères essentiels à
travers toutes les générations successives or cette fixité de l’espèce cesse
d’être un dogme à partir du moment où les observations des naturalistes
montrent qu’entre des espèces bien tranchées, peut exister un nombre
considérable de formes intermédiaires parmi lesquelles il est impossible
d’établir un point de séparation.
Une autre définition est alors proposée par Le Dantec : « ensemble de tous les individus qualitativement identiques et ne présentant entre eux, dans leurs
éléments vivants, que des différences quantitatives ».
En fait, la notion de
descendance reste le moyen de limiter et de définir à nouveau l’espèce, en
disant que tous les individus féconds entre eux et dont les descendants sont
indéfiniment féconds appartiennent à la même espèce.
Le travail de Robert Mazel a le mérite et l’intérêt
d’aiguiser notre sens de l’observation et notre sens critique pour nous guider
vers un consensus, tâche ardue pour les entomologistes que nous sommes.
Jacques
Comelade
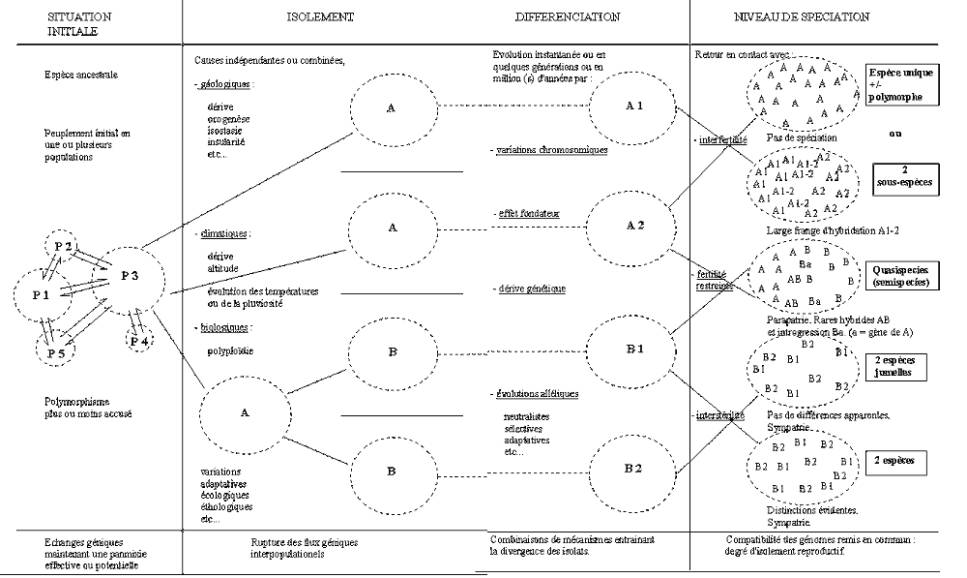
LA SPECIATION
SOMMAIRE (les
numéros correspondent aux numéros de pages de l’exemplaire papier)
RAPPEL DE QUELQUES NOTIONS DE RÉFÉRENCE. .............................................. 4
L'ADN, MOLÉCULE DU VIVANT. .................................................................................... 4
L'ADN, TOUT UN PROGRAMME.. ................................................................................... 6
LE CHANGEMENT : UNE ERREUR. ................................................................................. 6
LE BRASSAGE GÉNÉTIQUE. ............................................................................................ 7
LA GÉNÉTIQUE DES
POPULATIONS. ............................................................................. 7
LES FACTEURS
ENVIRONNEMENTAUX. ...................................................................... 8
ADAPTATION, ÉVOLUTION. ........................................................................................... 8
L'ISOLEMENT REPRODUCTEUR. .................................................................................... 8
ANALYSE DE CAS
CONCRETS RELEVÉS CHEZ LES LÉPIDOPTÈRES. ............... 9
FICHE 1
LE PEUPLEMENT D'EREBIA PANDROSE, " le Grand Nègre
bernois". ............................ 11
FICHE 2
LE PEUPLEMENT D'HELLEIA HELLE. .......................................................................... 12
FICHE 3
LE CONTACT ENTRE MELLICTA AURELIA ET MELLICTA PARTHENOIDES. ......... 13
FICHE 4
LE CONTACT ENTRE DEUX FORMES
DE ZYGAENA FILIPENDULAE. ................... 14
FICHE 5
L'ADAPTATION DIVERGENTE CHEZ
LES "DEMI-DEUIL". ......................................... 15
FICHE 6
ADAPTATIONS TROPHIQUES ET
ÉVOLUTION CHEZ LES "DAMIERS". .................. 16
FICHE 7
VARIATION DES PIÉCES GÉNITALES
CHEZ LA NOCTUELLE DE L'AUBÉPINE,
ALLOPHYES OXYACANTHAE L. ..................................................................................... 17
FICHE 8
STRUCTURATION DU GÉNOME. .................................................................................. 18
FICHE 9
DESCENDANCE D'UN MALE MUTANT
ROUX D'EUPHYDRYAS AURINIA.............. 19
FICHE 10
LE POLYMORPHISME
PHÉNOTYPIQUE AU SEIN DE L'ESPÈCE.
CHEZ L'ARGUS BLEU-NACRÉ (Polyommatus coridon). ............................................... 20
FICHE 11
POLYMORPHISME GÉNOTYPIQUE
INTRASPÉCIFIQUE. .......................................... 21
FICHE 12
POLYMORPHISME GÉNOTYPIQUE
INTERSPÉCIFIQUE. .......................................... 22
FICHE 13
UNE STASE ÉVOLUTIVE DANS LE
GENRE EUPHYDRYAS. ....................................... 23
LA SPÉCIATION DANS LE GROUPE
D'EREBIA TYNDARUS. ..................................... 24
L'ISOLEMENT GÉOGRAPHIQUE ET
SA RÉALISATION. ............................................ 26
1 - Le peuplement d'Erebia pandrose Borkhausen, le
"Grand Nègre bernois". (Fiche
1)
2 - Le peuplement d'Helleia helle Schiffermüller. (Fiche 2)
3 - Conclusion.
RETOUR EN CONTACT ET NIVEAU
DE SPÉCIATION. .............................................. 27
1 - Le contact entre Mellicta aurelia Nickerl et M. parthenoides Keferstein. (Fiche 3)
2 - Le contact entre deux
formes de Zygaena filipendulae L.. (Fiche 4)
ISOLEMENT ET DIFFÉRENCIATION
DES POPULATIONS. ....................................... 30
1 - L'adaptation divergente
chez les Melanargia ou
"Demi-deuil". (Fiche 5)
2 - Les adaptations
trophiques chez les "Damiers". (Fiche
6)
3 - Variations des pièces
génitales chez la Noctuelle de l'Aubépine, Allophyes
oxyacanthae L.
(Fiche 7)
ASPECTS GÉNÉTIQUES DE LA
SPÉCIATION. ............................................................. 34
1 - Structuration du génome.
(Fiche 8)
2 - Mutations. (Fiche 9)
3- Polymorphisme intra et inter-spécifique.
Le polymorphisme phénotypique au sein de l'espèce :
l'Argus bleu-nacré (Polyommatus
coridon, Poda). (Fiche 10)
Le polymorphisme génotypique intraspécifique. (Fiche 11)
Le polymorphisme génotypique interspécifique. (Fiche 12)
4 - Une stase évolutive dans
le genre Euphydryas. (Fiche 13)
SPÉCIATION COMPLEXE DANS LE
GENRE EREBIA. ................................................ 37
RÉFLEXION SUR
LA NOTION D'ESPÈCE ET SUR LA SPÉCIATION. ................. 38
LE CONCEPT DE L'ESPÉCE. ........................................................................................... 38
LE MODELE ALLOPATRIQUE DE LA
SPÉCIATION NÉODARWINIENNE. .............. 38
LA SPÉCIATION DANS LA
PRATIQUE ENTOMOLOGIQUE COURANTE. .............. 42
SPÉCIATION ET ÉVOLUTION. ....................................................................................... 44
ORIGINE DES
DOCUMENTS. ...................................................................................... 45
LA SPÉCIATION
ORIGINE ET
SÉPARATION DES ESPÈCES
Par Robert Mazel
En quelques décennies, les progrès de la biologie
moléculaire ont conduit à une interprétation cohérente des mécanismes
biologiques fondamentaux, particulièrement dans les domaines de la reproduction
sexuée et de la génétique. Comme tous les acquis, ceux-ci demeurent
perfectibles ou même discutables sur différents points mais les schémas de base
semblent définitivement établis : l'information génétique détenue par les
molécules d'ADN s'exprime en protéines par le truchement des ARN et elle est
transmise de génération en génération par les mécanismes de la méiose et de la
fécondation. La spéciation découle fondamentalement des modifications que subit
ce processus dans le temps et à l'échelle des populations.
L'approche
de ces phénomènes sera présentée ici en deux parties :
·
un
exposé posant un minimum de références théoriques fondamentales;
·
une
interprétation de cas réels présentés à partir de fiches.
En conclusion, sera proposée
une réflexion sur la notion d'espèce telle que la pratiquent quotidiennement
les entomologistes.
RAPPEL DE QUELQUES NOTIONS DE RÉFÉRENCE
L'ADN, MOLÉCULE DU VIVANT.
Une construction universelle de l'ADN est actuellement admise
: deux brins en hélice, formés de l'alternance de sucre, le désoxyribose, et
d'acide phosphorique, sont unis par les bases azotées que portent les sucres.
Il n'y a que quatre sortes de bases azotées unies deux à deux d'une seule
manière : Adénine avec Thymine et Guanine avec Cytosine. La séquence de ces
paires de bases, tout le long de la molécule, caractérise l'ADN d'une espèce
avec une faible variabilité d'un individu à un autre.
Chez l'homme, la longueur totale de l'ADN avoisine 2
mètres par noyau cellulaire et se répartit en 46 molécules. Celles-ci
constituent nos 46 chromosomes, ou mieux 23 paires : il existe en effet 2
molécules de chaque sorte dans toutes
les cellules dites diploïdes (symbole : 2n = 46), et un jeu simple de 23
chromosomes (= n) dans les gamètes, cellules haploïdes.
A différents moments de la vie cellulaire se produit une
véritable duplication de l'ADN, conduite par l'ADN-polymérase. Ce complexe
enzymatique rompt les liaisons entre bases azotées séparant ainsi
progressivement les deux brins de la molécule initiale et positionne
simultanément face à chacun de ces brins, de nouveaux nucléotides. Du fait de
la complémentarité univoque des bases azotées, A-T, C-G, il en résulte la
reconstitution de deux molécules d'ADN, rigoureusement identiques. Celles-ci
seront ultérieurement distribuées par une mitose, division cellulaire
équationnelle, en deux cellules filles.
Par ce moyen, toutes les cellules diploïdes d'un
organisme reçoivent une copie exacte et complète de l'ADN de la cellule-oeuf ou
zygote.
|
|
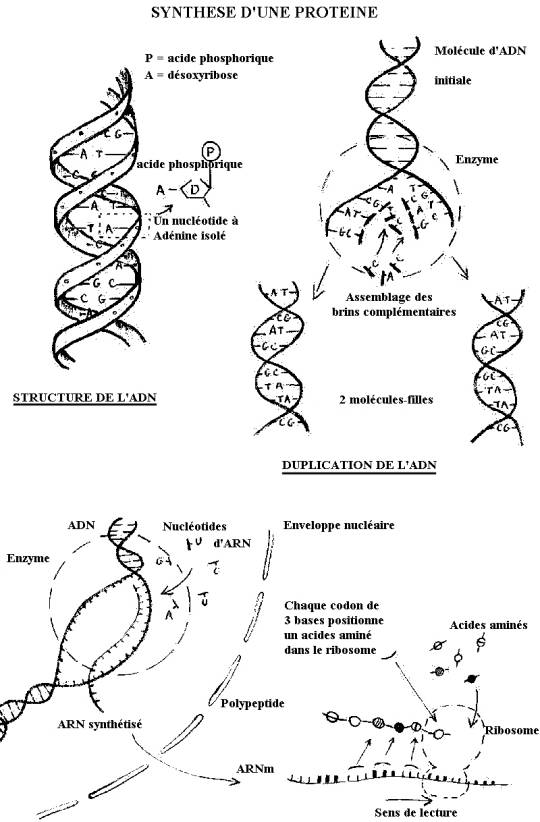
L'ADN, TOUT UN PROGRAMME.
En quoi ces molécules constituent-elles le patrimoine
génétique d'un individu ?
Un processus de transcription, voisin de la duplication,
permet de construire une molécule d'ARN à partie d'une région limitée de la
molécule d'ADN. Une ARN-synthétase intervient ici, qui positionne, en regard
d'un seul des brins de l'ADN, des nucléotides construits à partir de ribose,
Guanine, Cytosine, Adénine et Uracile. La molécule ainsi synthétisée,
complémentaire de la séquence d'ADN transcrite, constitue un simple brin qui se
sépare de l'ADN et passe dans le cytoplasme cellulaire. Il s'agit d'une
molécule d'ARN, de longueur variable mais sans commune mesure avec celle de
l'ADN.
Dans certaines régions du cytoplasme et par
l'intermédiaire de petits corpuscules, les ribosomes, l'ARN messager induit la
construction d'une chaîne d'acides aminés, c'est à dire d'un polypeptide qui
évolue directement en une protéine fonctionnelle.
Finalement, la séquence des bases azotées de l'ADN
détermine la succession des nucléotides de l'ARN qui sélectionne et positionne
à son tour les acides aminés de la molécule protidique. La portion d'ADN
permettant la synthèse d'un ARN messager est un gène, son expression est la
molécule protidique dont il gouverne la réalisation. En dernière analyse, le
message génétique n'est donc rien d'autre que l'ordre de succession des
nucléotides de l'ADN.
Les protéines synthétisées se répartissent en molécules
de structure, molécules de reconnaissance à l'origine des réactions
immunitaires, molécules fonctionnelles diverses : toutes les enzymes,
nombreuses hormones, neurotransmetteurs, substances de transport telles les
hémoglobines, de transfert ionique, etc. En d'autres termes, tous les
caractères d'un individu, morphologiques, anatomiques, physiologiques et
biologiques en général, sont fondamentalement déterminés par son ADN. Une
régulation complexe permet l'activation de tel ou tel gène selon les besoins
momentanés de l'organisme, le contrôle de son expression puis sa répression...
LE CHANGEMENT : UNE ERREUR.
Cependant, lors de la duplication de l'ADN, une ou
plusieurs bases azotées figurant dans la séquence initiale peuvent être
remplacées par d'autres. Cette erreur de copie constitue une mutation qui
produit une variante, un allèle nouveau, du gène initial. Si le caractère ainsi
modifié demeure viable, qu'il soit avantageux ou non, le nouveau gène sera
transmis comme l'ancien et augmentera donc le polymorphisme de l'espèce.
La fréquence des mutations, quoique très variable d'un
gène à l'autre, demeure très faible, de l'ordre de 10-9. Il n'en résulte pas moins
que les deux molécules d'ADN constituant une paire de chromosomes homologues
peuvent différer plus ou moins par les allèles qu'elles portent. Un individu
possédant deux allèles identiques d'un gène donné est dit homozygote pour le
caractère considéré, il est hétérozygote si les allèles sont dissemblables.
Dans ce cas, le caractère réalisé lors de l'expression de ce gène peut être
conforme à l'un des deux allèles ou intermédiaire : on parle respectivement de
dominance et récessivité ou absence de dominance... Il en résulte que tous les
gènes d'un individu, son génotype, ne s'expriment pas nécessairement dans son phénotype,
c'est à dire dans ses caractères apparents. Il s'agit là des bases élémentaires
de la génétique...
LE BRASSAGE GÉNÉTIQUE.
Mais la disparité des individus, en fait leur unicité, repose essentiellement sur la reproduction sexuée. Lors de la gamétogenèse, chaque cellule reproductrice ne reçoit que l'un des deux chromosomes de chaque paire de chromosomes homologues : les allèles se trouvent ainsi distribués au hasard avec les chromosomes qui les portent. La fécondation rétablit le nombre diploïde de chromosomes par juxtaposition dans le zygote des n chromosomes mâles et des n chromosomes femelles, également au hasard des rencontres entre gamètes. Ces processus entretiennent ainsi un brassage génétique continu des parents aux descendants.
Des échanges de segments de longueurs variables peuvent
en outre se produire entre molécules d'ADN homologues, lors de la méiose,
augmentant ainsi les recombinaisons alléliques. Toujours dans le déroulement de
la méiose, des erreurs dans la répartition des chromosomes conduisent à des
assemblages trisomiques ou à des zygotes triploïdes, tétraploïdes, etc...
Par essence, toute espèce apparaît donc nécessairement
polymorphe, à la fois dans l'espace et dans le temps.
LA
GÉNÉTIQUE DES POPULATIONS.
Une espèce comporte un nombre variable d'individus qui,
du fait de leur polymorphisme, détiennent chacun un certain assortiment des
allèles du génome commun, c'est à dire de la totalité des gènes qui définit une
espèce à un moment donné. Sur le terrain, ces individus constituent un
peuplement unique ou morcelé en populations plus ou moins isolées
géographiquement. Or les techniques d'investigation du génome, par séquençage
de l'ADN, par électrophorèse des protéines, etc., donnent accès à l'étude
statistique de la répartition de fréquence des allèles dans les populations. La
comparaison des distributions de fréquence allélique permet alors d'estimer le
degré d'éloignement des populations, de les rapporter à l'une ou l'autre de
deux espèces voisines, de vérifier la réalité de migrations géographiques,
etc... L'analyse en composants multivariables des phénotypes autorise également
une séparation fine qui recoupe les résultats précédants. (à moindre frais !).
A partir de ces données, divers indices d'identité génétique
ont été définis, fréquemment convertis en "distances génétiques", qui
peuvent traduire une certaine parenté phylétique. De manière générale, les
interprétations admettent un taux de mutation constant qui accroît
théoriquement l'hétérozygotie de manière infinie. Il existerait ainsi une
"horloge moléculaire" permettant de situer dans le temps les
mutations subies par une molécule donnée.
Par ailleurs, les mitochondries, organites cytoplasmiques
intervenant dans la production d'énergie cellulaire, possèdent également un peu
d'ADN. Seules les mitochondries de l'ovule se retrouvent dans le zygote de
sorte que l'analyse de l'ADN mitochondrial permet une "recherche en
maternité" dans les parentés phylétiques.
Sur le terrain, les techniques de capture-recapture et de
nombreuses autres méthodes d'analyse des effectifs, à formulation mathématique
plus ou moins complexe et subtile, tentent de rendre compte des fluctuations et
des migrations populationnelles. La notion de métapopulation, introduite depuis
peu, correspond à un peuplement plus ou moins morcelé ou à un ensemble de
populations qui évoluent conjointement par suite de migrations et d'échanges
géniques entre elles. Généralement une population principale peut être
distinguée de populations satellites. Un dispositif de ce type, assez dispersé,
peut pallier des extinctions localisées par repeuplement à partir d'une
population mère...
LES
FACTEURS ENVIRONNEMENTAUX.
Ils
se répartissent en facteurs :
-
climatiques,
-
édaphiques
et biotiques, incluant les actions
anthropiques. Il s'agit là du domaine écologique.
A
l'échelle du globe, les variations latitudinales et altitudinales du climat
rendent compte de la zonation et de l'étagement des peuplements, aussi bien
végétaux qu'animaux. Mais en fait, les microclimats régnant sous une écorce,
dans une fissure du sol ou sous une touffe de végétation ont généralement une
plus grande incidence dans la localisation d'une majorité d'insectes que le
climat général. La relation s'établit ainsi avec les caractéristiques
édaphiques et les espèces se répartissent selon leurs preferenda.
L'organisation de détail des peuplements découle, le plus fréquemment, des
interactions biotiques inter- et intraspécifique : relations trophiques,
prédation, parasitisme, compétition, comportements reproducteurs, etc...
L'Homme interagit à tous les niveaux, le plus souvent en modifiant les
conditions originelles.
L'organisation
des biocénoses, communautés vivantes propres à un biotope donné, résulte ainsi
de toutes les interactions qui s'exercent. Dans un milieu relativement stable
s'établit un équilibre fluctuant, souvent cyclique ou périodique, qui répond à
toute perturbation durable en se transformant et en évoluant vers un nouvel
équilibre.
ADAPTATION,
ÉVOLUTION.
Du fait de leur polymorphisme, tous les individus d'une
espèce ne sont pas aptes également à subir telle ou telle contrainte de leur
environnement. Ainsi les mutations peuvent faire apparaître un allèle
favorable, c'est à dire conférant à celui qui le porte une meilleure compétitivité
dans une situation donnée. Il en résultera la transmission et la conservation
de cet allèle dans les générations suivantes. L'avantage sera en quelque sorte
sélectionné par le milieu et participera à l'adaptation-évolution de l'espèce.
En d'autres termes, les espèces se transforment nécessairement dans l'espace et
dans le temps par suite des pressions qu'exercent les conditions de vie sur les
individus.
Cependant et en conséquence de leur caractère aléatoire
et imprévisible, les mutations ne confèrent aucune orientation ni aucune valeur
intrinsèque aux modifications phénotypiques qu'elles induisent. Elles sont
neutres au regard de l'évolution et ce sont les pressions de sélection qui
orientent le processus dans un sens apparaissant favorable sur une période plus
ou moins longue. Ainsi se différencie la majeure partie des sous-espèces
géographiques en réponse aux variations environnementales que rencontrent les
populations d'une espèce dans son aire de dispersion.
Dans le temps, deux populations de la même espèce vont
progressivement diverger par orientation de la sélection dans des directions
différentes. Ce processus peut-il alors aboutir à séparer deux nouvelles
espèces ?
L'ISOLEMENT REPRODUCTEUR.
Quels que soient ses attributs par ailleurs, une espèce
constitue d'abord et obligatoirement une communauté d'individus interfertiles.
Tant que deux populations cospécifiques demeurent en contact, même par un
espace réduit ou périodiquement et de manière aléatoire, le brassage
chromosomique entretient l'échange de gènes et maintient la cohésion de
l'espèce.
A contrario, la séparation de deux espèces issues d'une
même peuplement initial implique l'instauration d'une barrière d'inter
stérilité. Le critère fondamental de la spéciation est donc l'isolement
reproducteur entre deux communautés.
Si la logique de ces principes demeure incontestable, il
n'en va pas de même de la discussion des innombrables questions qui surgissent
dès que l'on se réfère aux faits observés sur le terrain. En particulier,
l'isolement reproductif apparaît-il en conséquence de la divergence évolutive
ou en est-il une composante originelle ? Ce sont donc les données de
l'observation et de l'expérimentation qu'il convient d'analyser avant de proposer
une conclusion.
ANALYSE DE CAS CONCRETS
RELEVÉS CHEZ LES LÉPIDOPTÈRES.
Les documents présentés dans les fiches qui suivent,
directement issus du terrain ou de recherches diverses, illustrent les aspects
essentiels ou les plus fréquents de spéciation conformes au schéma néodarwinien
proposé en conclusion. Cependant le choix des données reste volontairement
limité aux domaines de connaissances et de réflexion accessibles au plus grand
nombre, sensiblement au niveau des classes terminales de lycée.
La démarche fondamentale repose sur l'analyse de données
expérimentales dont la richesse demeure préservée par une présentation aussi
neutre que possible, réduite à un titre accompagné éventuellement de quelques
indications complémentaires facilitant leur seule compréhension. Ces documents
sont exploités en une série de questions centrées sur la spéciation, ou l'un de
ses mécanismes, et formulées sous la forme de "problèmes posés". Une
interprétation des données, parfois enrichie de quelques commentaires plus
généraux, répond directement à ces questions.
Enfin, un texte de liaison fournit des informations
supplémentaires et des notes diverses qui élargissent certaines études ou les
replacent dans la complexité de leur cadre d'origine.
Les espèces étudiées sont
présentées et figurées ci-après, eu quasi-totalité, dans la reproduction d’une
planche due au talent de Christian
Corraze.
(Photos dans le document papier)
1 - Erebia pandrose
(Le Grand Nègre bernois). Espèce adaptée aux climats boréo-alpins comme
beaucoup d’Erebia.
2 - E.
hispania; 3 - E.
cassioïdes. Deux des espèces du groupe tyndarus, ancienne espèce collective actuellement éclatée en 7
taxons, au moins, répartis dans les massifs européens. 4 - Helleia helle. Lycène caractéristique de la faune des
tourbières froides. 5 - Polyommatus
bellargus (Azuré bleu céleste). 6
- P.
coridon (Argus bleu nacré). Le femelle est brune comme celle de bellargus.
Les deux espèces s’hybrident en dépit de formules chromosomiques très
différentes. 7-8-9 - Zygaena
filipendulae.
L’une des Zygènes les plus communes de France. Coloration aposématique en
rapport avec son caractère immangeable et toxique pour les oiseaux. 10 - Mellicta parthenoïdes; 11 - M. aurelia. Deux Mélitées
voisines, en partie sympatriques en France, mais d’origines biogéopgraphiques
distinctes. 12 - Euphydryas
iduna. Aire atypique : nord de la Scandinavie, Caucase, monts Altaï et
Sajan. Quelques espèces voisines des Alpes, une des Montagnes Rocheuses
(Montana). Stases évolutives ? 13 - Euphydryas
aurinia pyrenes-debilis, au-dessus de 2000 m dans les
Pyrénées-Orientales; 14 - E.
aurinia beckeri, basse altitude en Espagne. Contrairement au groupe
précédent d’E. iduna, E. aurinia présente un foisonnement de
sous-espèces de l’Irlande au Japon. 15
- E.
aurinia, mutation rousse.
L’élevage ayant été arrêté, cet animal n’existe plus... 16 - E. editha. Espèce d’Amérique du Nord présentant une extension
comparable à celle d’E. aurinia en
Eurasie. 17 - Melanargia galathea (Le
Demi-Deuil). Commun dans toute la France. Ses populations du sud-est (18) présentent une forte mélanisation.
FICHE 1 LE PEUPLEMENT D'EREBIA
PANDROSE, " le Grand Nègre bernois".
|
|
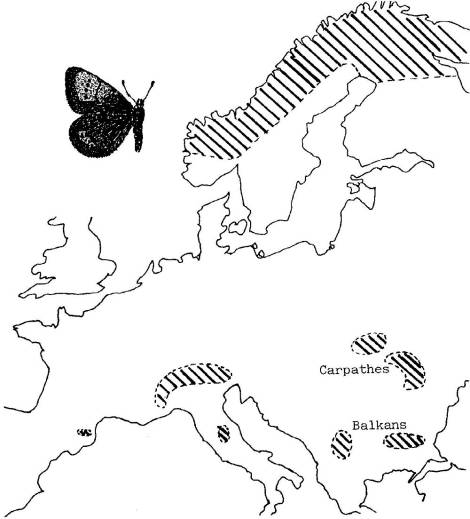
Carte de répartition
européenne.
L'espèce est présente dès le niveau de la mer dans
le nord de la Scandinavie et au-dessus de 1800 m dans les Alpes et les
Pyrénées.
Les chenilles, peu exigeantes, se nourrissent de
plusieurs espèces de Graminées.
Problèmes
posés :
- Comment s'explique la localisation des populations
de l'espèce ?
- Comment a pu se réaliser ce peuplement ?
FICHE 2 LE PEUPLEMENT D'HELLEIA
HELLE.
|
|
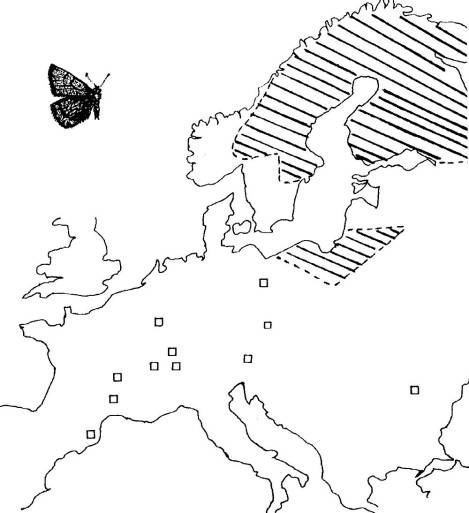
Carte de répartition.
Hachures
: peuplement continu au nord de l'Europe.
Carrés
: populations isolées.
Helleia helle vit dans les zones humides
des régions froides où poussent la Bistorte (Polygonum bistorta) et
la Patience (Rumex
sp.) dont se nourrissent ses chenilles. Le papillon ne s'éloigne guère de ces
mêmes biotopes.
Problèmes
posés :
-
Comment peut s'expliquer la distribution de H.
helle ?
-
Quelles hypothèses suggère-t-elle quant à son origine ?
FICHE 3 LE CONTACT ENTRE MELLICTA
AURELIA ET MELLICTA PARTHENOIDES.
|
|
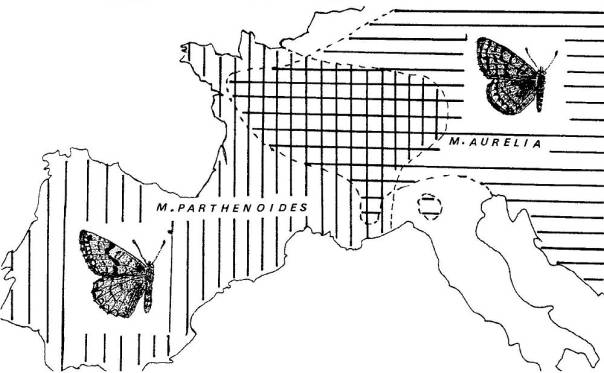
Ces deux
Mélitées possèdent de nombreux caractères communs ou très proches, à tel point
que leur appartenance à une seule espèce ou à deux espèces distinctes pourrait
être discutée.
Aires biogéographiques des deux taxons.
Mellicta aurelia s'établit essentiellement dans les prairies
naturelles, jusque vers 1500 m d'altitude. Ses chenilles se nourrissent de
Plantain, Véronique et Mélampyre.
Mellicta parthenoides fréquente les prairies, les
zones découvertes, les bordures de bois, où croissent les plantes-hôtes de ses
chenilles, en particulier Plantain, Mélampyre et Scabieuse. Dans les Pyrénées,
elle se trouve jusqu'à plus de 2000 m.
Les deux
espèces cohabitent fréquemment mais on n'observe pas d'hybride dans leur zone
de sympatrie.
Problèmes
posés :
-
La biologie de ces deux Mélitées explique-t-elle leur répartition géographique
?
-
Comment celle-ci a-t-elle pu se réaliser ?
-
S'agit-il, en fait, d'une ou de deux espèces ?
FICHE 4 LE CONTACT ENTRE DEUX FORMES DE ZYGAENA FILIPENDULAE.
|
|
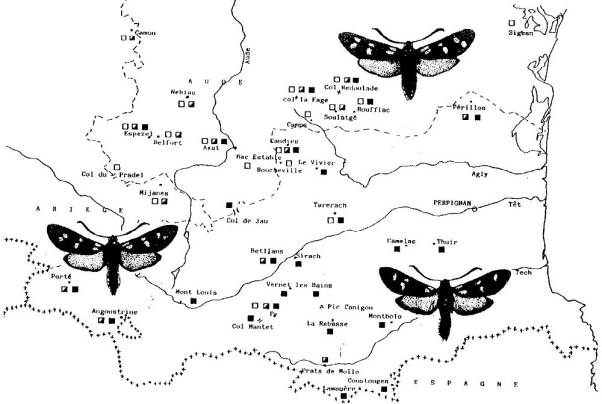
La Zygène de
la Filipendule, largement répandue en Europe, s'orne de six taches rouges sur l'aile antérieure. Cependant, des
peuplements à cinq macules existent en Espagne, dans le Sud de la France et en
Italie.
· Répartition de la Zygène de la Filipendule (Zygaena filipendulae) de l'Aude aux Pyrénées-Orientales.
Formes à 5 macules (carré noir), à 6 macules (carré blanc) et intermédiaires? Tout le peuplement espagnol frontalier est à 5 taches.
En élevage,
les formes à 5 ou 6 macules se sont révélées pures; en revanche, des individus
à 6 taches, 5 taches "un quart" et 5 taches ont été obtenus
simultanément de la ponte d'une femelle à 5 taches "et demi".
Problèmes posés :
- Quel est le palier de
spéciation atteint par ces différentes Zygènes ?
- Comment s'explique leur
répartition ?
FICHE 5 L'ADAPTATION DIVERGENTE CHEZ LES
"DEMI-DEUIL".
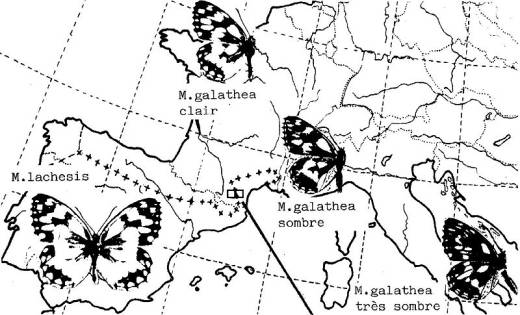
Selon les auteurs, Melanargia lachesis est considéré comme sous-espèce de M. galathea ou comme espèce distincte.
I -
Peuplements des deux Melanargia.
|
|
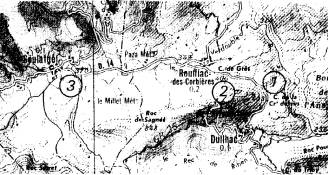
II - Etude d'une zone de contact dans
les Corbières.
|
Composition
de 3 échantillons observés le
même jour : 1 - Col de
la Croix dessus : 30 M.
lachesis; 2 - Col de
Grès : 12 lachesis 14
galathea, 7 hybrides; 3 - Soulatgé : 35 galathea. |
Problèmes
posés :
- Pourquoi parle-t-on d'adaptation divergente ?
- Comparer la proportion d'hybrides et l'extension
de la frange d'hybridation
à celles observées
chez Z. filipendulae.
- Comment comprendre que les deux peuplements ne se
mêlent pas ou ne se confondent pas ?
Faut-il conclure à l'existence de deux
espèces distinctes ou d'une seule ?
FICHE 6 ADAPTATIONS TROPHIQUES ET ÉVOLUTION CHEZ LES
"DAMIERS".
Le groupe des
Damiers, Euphydryas et Melitaea au sens large, compte une
trentaine d'espèces en Eurasie et quelques unes en Amérique du Nord. Chaque
espèce pond sur une ou quelques espèces végétales nourricières des chenilles,
toutes apparentées botaniquement et chimiquement par les composés qu'elles renferment.
En France, les sous-espèces du Damier de la Succise, Euphydryas aurinia, illustrent à elles seules cette spécificité
trophique.
|
Sous-espèces d' E. aurinia
et leurs plantes-hôtes, en France. |
||
|
Ssp.
d'E. aurinia |
Biogéographie |
Plantes-hôtes |
|
aurinia aurinia aurinia xeraurinia aurinia provincialis aurinia beckeri aurinia pyrenes-debilis aurinia glaciegenita |
Zones
humides. 0 à 1800 m env. Coteaux
±Erreur! Signet non défini. secs du sud Sud-est
à basse altitude Pyr.Or.
à basse altitude Pyr.Or.
de 1500 à 2500 m Alpes
vers 2000 m et plus |
Succise Scabieuse,
Knautie Céphalaire Chèvrefeuille Succise
puis Gentiane alpine après 2000 m. Gentiane de Koch |
|
Elevage d'hybrides sur les plantes-hôtes parentales. |
|
Parents Plante d'élevage des
chenilles Nbre
d'imagos |
|
™ provincialis X = 1 ponte ÚErreur! Signet non défini. Chèvrefeuille
.................................. 133 ˜Erreur! Signet non défini. beckeri ™Erreur! Signet non défini. beckeri
ÛErreur! Signet non défini. Chèvrefeuille
................................. 162 X = 2 pontes ˜Erreur! Signet non défini. provincialis
ÙErreur! Signet non défini. Céphalaire
......................................... 3 ™Erreur! Signet non défini. provincialis ÛErreur! Signet non défini. Erreur! Signet non défini. Céphalaire ..................................... 134 X = 2 pontes ˜Erreur! Signet non défini. aurinia ÙErreur! Signet non défini. Succise ..............................................
0 ™Erreur! Signet non défini. aurinia ÛErreur! Signet non défini. Erreur! Signet non défini. Céphalaire ....................................... 36 X = 4 pontes ˜Erreur! Signet non défini. provincialis ÙErreur! Signet non défini. Succise
.............................................. 2 |
Problèmes posés :
- Quelle est la nature de la
spécificité trophique observée ?
- Quelles hypothèses expliqueraient
la diversification des sous-espèces d'E. aurinia et,
par analogie, la spéciation dans ce groupe de Lépidoptères ?
FICHE 7 VARIATION DES PIÈCES GÉNITALES CHEZ LA NOCTUELLE DE
L'AUBÉPINE,
ALLOPHYES OXYACANTHAE L.
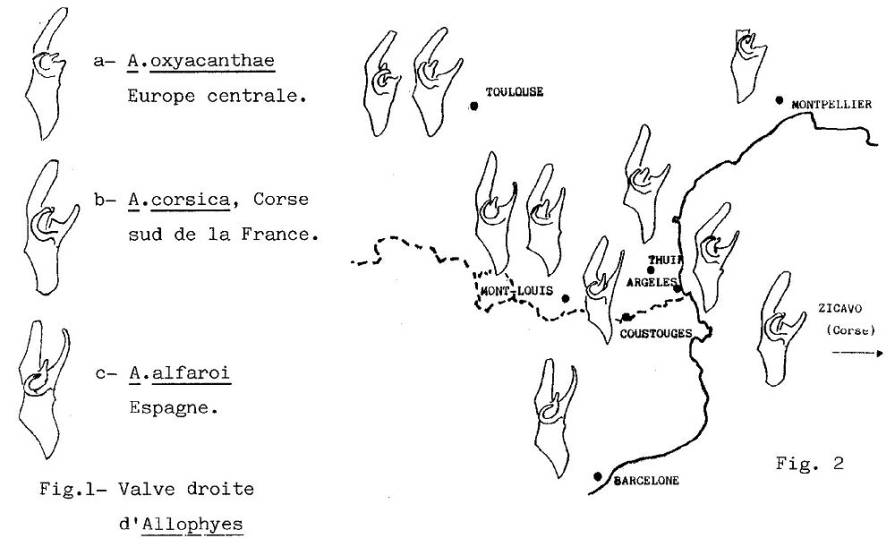
Cette
Noctuelle, commune en Europe, présente partout la même ornementation bariolée qui la rend très peu visible au
repos sur les rochers, les troncs, etc... Les mâles portent des appendices
abdominaux en pinces, les valves, qui maintiennent la femelle pendant
l'accouplement et on a constaté que la conformation de ces pièces chez les Allophyes de la péninsule ibérique
diffère fortement de celle connue partout ailleurs en Europe continentale.
(Fig. 1). On a ainsi séparé l'espèce alfaroi
Ag., d'A. oxyacanthae L. La
découverte d'une troisième conformation en Corse, a justifié de même la
description d'A. corsica Spuler.
(Fig. 1b.). Des études récentes remettent en question ces séparations en
établissant l'existence de structures de type corsica dans le sud de la France. De plus, ces caractères se
recombinent indépendamment à d'autres caractères d'alfaroi et d'oxyacanthae
dans les populations du Roussillon, de la région toulousaine, etc... (Fig. 2
ci-dessous).
Problèmes
posés :
-
Comment s'explique le polymorphisme des valves des mâles par rapport à la
stabilité de l'ornementation de ces papillons ?
- Quels arguments permettent d'affirmer que la structure génétique de l'ensemble du peuplement européen est très probablement celle d'une espèce unique ?
FICHE 8 STRUCTURATION DU GÉNOME
Chez les
Lépidoptères, les chromosomes appartiennent généralement au type holocentrique,
de forme plus ou moins globuleuse. Leur nombre moyen et le plus fréquent est n
= 30. Chez les "Petits bleus" du genre Polyommatus, au sens large, on observe les formules chromosomiques
ci-dessous, en métaphase haploïde.
|
|

1 - Polyommatus syriaca
2 - P. zuleika
3 - P. bellargus
4 - P.
coridon
5
et 6 - Hybrides coridon X bellargus
7 - P.
golgus
8 - P.
agester
9 -
P. atlantica
(Probablement le nombre de
chromosomes le plus élevé connu chez les Animaux).
Problèmes posés :
- Comment comprendre une
telle diversité chez ces espèces relativement voisines ? Quelles conséquences évolutives peuvent
avoir ces variations ?
-
Les formules 5 et 6 ont été relevées chez des hybrides naturels, P. coridon X
P. bellargus, fertiles sur une ou deux générations. Que peut-on en déduire ?
FICHE 9 DESCENDANCE D'UN MALE MUTANT ROUX D'EUPHYDRYAS AURINIA.
Un mâle totalement roux ferrugineux, apparu dans un
élevage en 1983, s'est accouplé avec une femelle normalement ornementée. Le
caractère roux a été suivi pendant 4 ans au travers de croisements
partiellement rapportés ici. La mutation n'a, semble-t-il, jamais été observée
dans la nature. Les mutants ont montré, en moyenne, une durée de vie courte et
une mauvaise résistance à l'exposition au soleil.
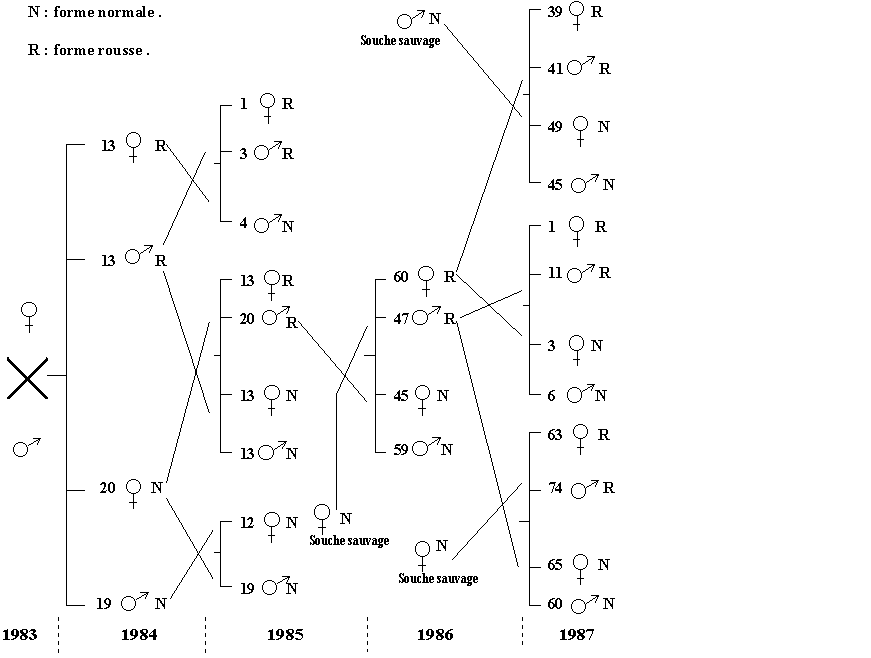
Croisement
™ normale X ˜ roux.
Problèmes
posés :
-
Déterminer le mode de dominance du gène muté. Est-il autosomique ou lié aux
chromosomes sexuels ?
-
Etablir les génotypes des parents et descendants de ces croisements.
-
Quelle peut être l'incidence de ce type de mutation dans la spéciation ?
FICHE 10 LE POLYMORPHISME PHÉNOTYPIQUE AU SEIN DE L'ESPÈCE
CHEZ L'ARGUS
BLEU-NACRÉ (Polyommatus coridon).
|
|
Chez les "Petits-Bleus" ou "Argus", seul le mâle est ordinairement bleu, la femelle étant brune; cependant des femelles bleues existent dans certaines populations.
D'autres
populations à fort pourcentage de femelles bleues, jusqu'à 100%,
sont connues
en Espagne et en Italie.
Problèmes
posés :
- La
répartition des femelles bleues peut-elle être corrélée avec des facteurs environnementaux,
géographiques, climatiques, etc, ou doit-elle être considérée neutre vis à vis
de ces facteurs ?
- Un tel
polymorphisme représente-t-il un avantage évolutif pour l'espèce considérée ?
Remarque : on
a démontré que bleu est récessif par rapport à brun.
FICHE 11 POLYMORPHISME GÉNOTYPIQUE
INTRASPÉCIFIQUE.
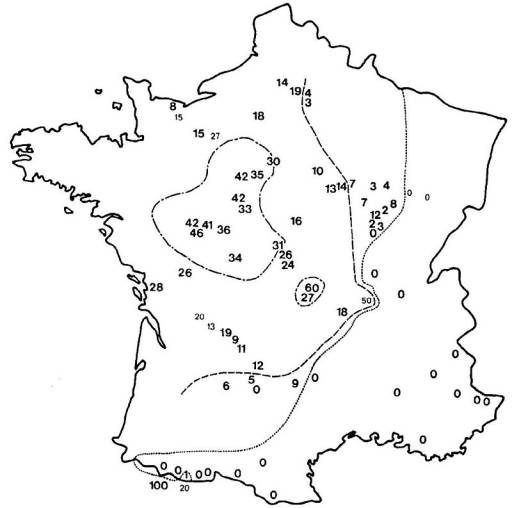
L'activité des organismes est dirigée par les enzymes du métabolisme général, intracellulaires, elles-mêmes produites par les gènes qui les contrôlent. Pour un gène pluriallélique, les individus d'une même population peuvent synthétiser des molécules enzymatiques différentes, séparables par migration dans un champ électrique selon leurs charges et leurs masses, c'est à dire par électrophorèse. On distingue ainsi les allèles présents à un locus donné et on établit les génotypes d'une population pour un ou plusieurs gènes. Certains gènes se révèlent strictement monomorphes si tous les individus, homozygotes, ne portent qu'un seul et même allèle. D'autres gènes, polymorphes, sont représentés par plusieurs allèles qui occupent le locus considéré à des fréquences différentes.
Les fréquences de 5 allèles codant pour la
phosphoglucomutase (PGM) ont ainsi été établies, en pourcentage pour 20 mâles,
dans diverses populations de Lysandra hispana
H.S. du sud de la France et du nord de l'Espagne.
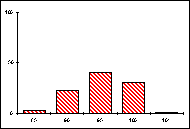
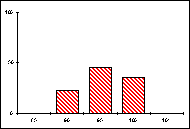
Guera : Aix-en-Diois Lauroux Uzés
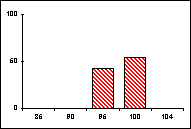
Barjols Montllobat Atares
L'allèle le plus fréquent est noté 100, les autres
sont repérés en fonction de leur écart mesuré en cm sur le gel
d'électrophorèse.
Problèmes
posés :
-
Quelles informations apporte ce type d'études sur la structure génotypique
d'une espèce ?
-
Comment comprendre les disparités mises en évidence ?
-
Quel intérêt ou quel risque cela constitue-t-il pour l'espèce ?
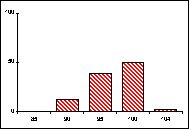
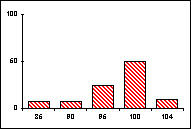
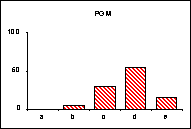
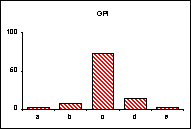
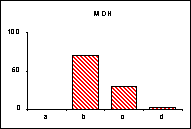
FICHE 12 POLYMORPHISME GÉNOTYPIQUE INTERSPÉCIFIQUE.
Dans les Hautes-Alpes, près de
Châteauneuf-de-Châbre, volent ensemble 3 espèces voisines : Lysandra (=Polyommatus) hispana
(H.S.), L. coridon (Poda) et L. bellargus (Rott.) Les fréquences
alléliques ont été déterminées, en pourcentage sur des échantillons de 20
mâles, pour 3 gènes codant les enzymes : PGM, phosphoglucomutase, GPI, glucose
phosphate isomérase, MDH, malate déshydrogénase.
Lysandra hispana
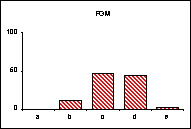
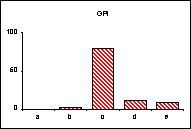
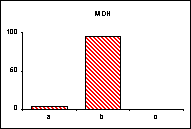
Lysandra coridon
Lysandra bellargus
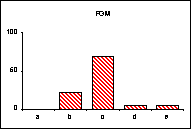
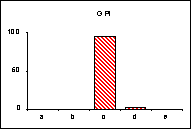
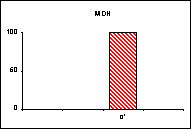
L'allèle
c' de la MDH n'a été trouvé ni chez L.
hispana ni chez L. coridon.
Problèmes
posés :
-
Quelles caractéristiques présente la variation allélique comparée chez ces
trois espèces ?
-
Quelles conséquences adaptatives et évolutives peuvent en découler ?
-
Pour l'étude des populations et de la spéciation, quels intérêts offre la
méthode utilisée ?
FICHE 13 UNE STASE ÉVOLUTIVE DANS LE GENRE EUPHYDRYAS.
|
|
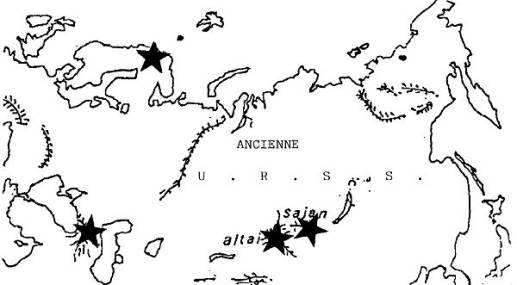
Répartition d'Euphydryas iduna Dalman.
D'une station à l'autre, E. iduna ne présente pas de variations
morphologiques importantes.
Quatre autres espèces voisines, mais bien tranchées,
se localisent :
- dans les Alpes : E. intermedia;
- dans les Alpes et les Balkans : E. cynthia;
- au nord de l'Europe et en Russie : E. maturna;
- dans les montagnes Rocheuses du Montana : E. gillettii.
Ces espèces paraissent
toutes remarquablement stables, bien qu'elles constituent généralement de
petits peuplements épars.
Problèmes posés :
- En quoi les observations faites chez ces Euphydryas semblent elles remettre en question
les mécanismes évolutifs ?
- Quelles hypothèses pourraient expliquer
la stabilité observée ?
LA SPÉCIATION DANS LE GROUPE
D'EREBIA TYNDARUS.
|
|
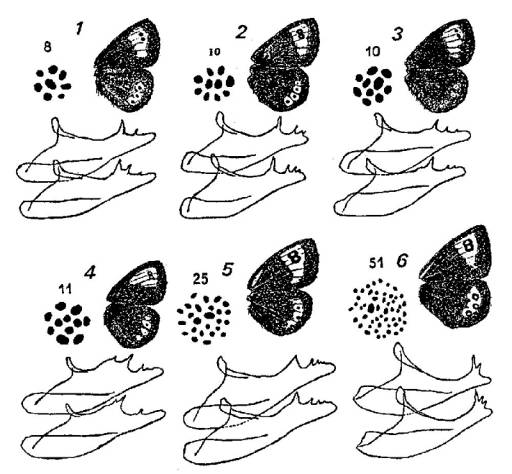
Ornementation alaire
d'espèces voisines du genre Erebia, caryotypes
(formules haploïdes = n chromosomes) et forme des valves de l'appareil
copulatoire des mâles. (Ces pièces maintiennent l'abdomen des femelles pendant
l'accouplement).
1 - E. calcarius; 2 - E. cassioides; 3 - E. tyndarus;
4 - E. nivalis; 5 - E. hispania; 6 - E. iranica.
Cette dernière espèce habite certaines zones
subalpines d'Asie Mineure.
Données complémentaires :
Dans les Pyrénées, cassioides et hispania entrent en contact sans hybridation mais s'excluent
réciproquement d'un même biotope.
Dans les Alpes, les mêmes
relations s'établissent entre nivalis
et tyndarus ou nivalis et cassioides.
Les
hybridations expérimentales réalisées entre plusieurs de ces espèces ont révélé
la stérilité des hybrides obtenus. La forme des valves ne constitue pas un
obstacle à ces accouplements.
Répartition
géographique des espèces du groupe d'Erebia
tyndarus en Europe.
|
|
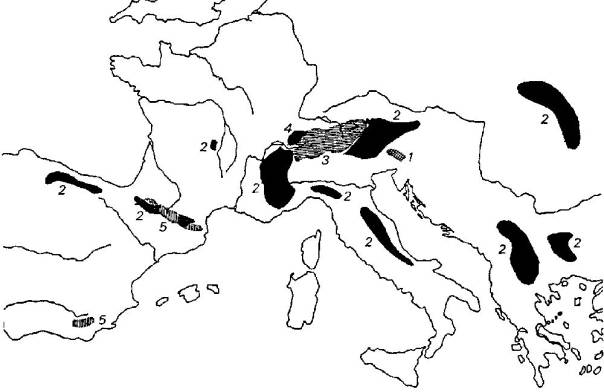
1 - Erebia calcarius 2 - Erebia cassioides
3 -
Erebia tyndarus 4 - Erebia nivalis
5 - Erebia
hispania
Une trentaine d'autres
espèces appartenant au genre Erebia,
toutes très sombres ou noires, peuplent les mêmes biotopes d'altitude. Le
groupe d'E. tyndarus constitue un complexe spécifique homogène qui tranche
parmi les autres Erebia plus
diversifiés.
Comment peut-on concevoir la
spéciation dans ce groupe de Lépidoptères gravitant autour d'Erebia tyndarus ?
Une équipe de chercheurs
italiens a présenté en 1992 les résultats d’électrophorèses d’enzymes réalisées
chez ces mêmes Erebia.
L’interprétation des analyses conduit à séparer deux groupes de populations au
sein d’E. cassioides : le peuplement
des Alpes Centrales et Orientales opposé à l’ensemble Occidental occupant les
Apennins, l’ouest des Alpes, les Pyrénées et les Monts Cantabriques. Les
distances génétiques entre les deux groupes apparaissent significatives bien
que relativement faibles.
Comment ces nouvelles
données s’intègrent-elles aux précédentes ?
L'ISOLEMENT GÉOGRAPHIQUE ET SA RÉALISATION.
1 - Le
peuplement d'Erebia pandrose Borkhausen, le "Grand
Nègre bernois". (Fiche 1)
Interprétation:
Les
populations actuelles se localisent dans l'extrême nord européen ou en altitude
sur les reliefs. Les chenilles n'étant pas strictement tributaires d'une espèce
végétale, ce sont vraisemblablement les facteurs climatiques qui déterminent
cette répartition : l'espèce apparaît liée à des milieux froids.
Remarque : les mêmes exigences
biologiques se retrouvent chez les autres Erebia,
plus de 30 espèces européennes boréo-alpines ou endémiques d'altitude, toutes
de coloration très sombre ou totalement noires. Il s'agit donc de
caractéristiques communes à tout le genre Erebia.
Le
morcellement de l'aire d'E. pandrose traduit la réponse de l'espèce
à des fluctuations climatiques récentes, en rapport avec les glaciations
quaternaires. Le scénario le plus simple doit comporter une première séquence
d'expansion : l'espèce adaptée aux climats froids, favorisée par un ou
plusieurs refroidissements, s'étend vers le sud. Le réchauffement relatif
actuel entraîne, à l'inverse, un reflux en Scandinavie ou en altitude. Les
populations réfugiées sur les sommets constituent alors de véritables
peuplements insulaires piégés par la variation climatique.
2 - Le
peuplement d'Helleia helle Schiffermüller. (Fiche 2)
Interprétation
:
La
distribution européenne de l'espèce se rapproche de celle d'Erebia pandrose. Cependant les
populations isolées ne semblent pas strictement liées aux climats d'altitude
mais plutôt aux biotopes froids et suffisamment humides qui permettent le développement
des plantes-hôtes de l'espèce. La répartition actuelle résulterait ainsi de
l'extension des zones humides sous un climat froid puis d'une régression ne
laissant subsister que quelques populations dans des biotopes résiduels liés à
des situations stationnelles particulières.
L'isolement
résulte donc ici de la combinaison de facteurs climatiques, édaphiques et
trophiques pour les larves.
Remarque : Ces populations constituent
actuellement des reliques subglaciaires associées à un cortège floristique
caractéristique des tourbières, très fragile.
L'étude
d'Helleia helle peut être complétée
comme suit :
les
populations de l'est de la France, du Massif Central, des Pyrénées, etc, ont
été séparées en sous-espèces différant par certains caractères morphologiques
des papillons. Comment s'expliquent de telles différences ?
Les
variations morphologiques relevées traduisent évidemment la dérive génétique et
les pressions de sélection différentes s'exerçant sur chaque isolat.
3 -
Conclusion.
Les
fluctuations du climat peuvent suffire à produire le morcellement et
l'isolement géographique des populations liées directement ou indirectement aux
facteurs climatiques par leurs exigences biologiques.
La
colonisation d'un archipel par des espèces migratrices ou introduites
fortuitement conduit à des situations analogues de même que les manifestations
tectoniques de dérive, orogenèse, etc...
Les
être vivants peuvent encore générer eux-mêmes les conditions de leur
ségrégation en sous-ensembles isolés par leur comportement ou par des
modifications chromosomiques dont ils sont le siège. Des exemples d'isolements
comportementaux ou chromosomiques sont donnés plus loin par l'étude des Euphydryas et des Lycènes. Fiches 6 et
8.
RETOUR EN
CONTACT ET NIVEAU DE SPÉCIATION
1 - Le contact
entre Mellicta aurelia Nickerl et M. parthenoides Keferstein. (Fiche 3)
L'appartenance
au même genre et le graphisme très voisin des ailes montrent la très proche
parenté de ces deux Mellicta. De plus
elles fréquentent les mêmes biotopes et leurs plantes-hôtes sont communes ou
apparentées de sorte qu'il est impossible de pronostiquer leur appartenance à
une ou deux espèces à partir de ces seuls critères. En revanche, leur
cohabitation sans échange génique témoigne immédiatement de leur séparation
spécifique.
Les
situations biogéographiques actuelles résultent de la dynamique postglaciaire
des peuplements. La péninsule ibérique a servi de refuge à M. parthenoides de même
qu'à l'ensemble des espèces refoulées vers le sud-ouest de la France par
l'extension de l'inlandsis polaire. Cependant la séparation spécifique des Mellicta aurelia et parthenoides, relativement bien tranchée, remonte à une époque
certainement antérieure au dernier épisode glaciaire.
2 - Le contact
entre deux formes de Zygaena filipendulae
L. (Fiche 4)
Interprétation
:
Les
formes à "5 taches ½" apparaissent au contact du peuplement espagnol
à 5 taches et des formes à 6 taches, traduisant le brassage de ces caractères
sur des dizaines de km. De plus, les données expérimentales établissent la
fertilité de ces formes intermédiaires. Un échange génique sans restriction se
réalise ainsi entre populations à 6 et 5 macules qui ne représentent donc, au
plus, que des sous-espèces d'une même espèce.
Les
populations à 5 macules apparaissent confinées aux régions les plus chaudes de
l'aire de l'espèce, méditerranéennes en particulier; elles pourraient donc
représenter une forme mieux adaptée à ces contraintes climatiques et
inversement pour le type à 6 taches. Le caractère marquerait alors une
divergence adaptative, celle-ci ne s'accompagnant pas d'une séparation
spécifique. Le retour en contact, vraisemblablement provoqué ici par le
réchauffement climatique, a alors pour conséquence une augmentation du
polymorphisme de l'espèce, traduite notamment par la réalisation des formes
intermédiaires.
3 - Documents
complémentaires.
Une
analyse plus large du peuplement de Zygaena
filipendulae a été effectuée, notamment en France. Des formes à 5 et 6
macules viennent également en contact dans le sud-est : la carte établie pour
cette région peut-être comparée à celle obtenue dans les Pyrénées-Orientales.
Enfin, les proportions des deux formes dans quelques stations de l’Aude, de
l’Aveyron, de l’Hérault, etc montrent les rapports établis avec le peuplement
est-pyrénéen.
A - Peuplement dans le sud-est de la France.
|
|
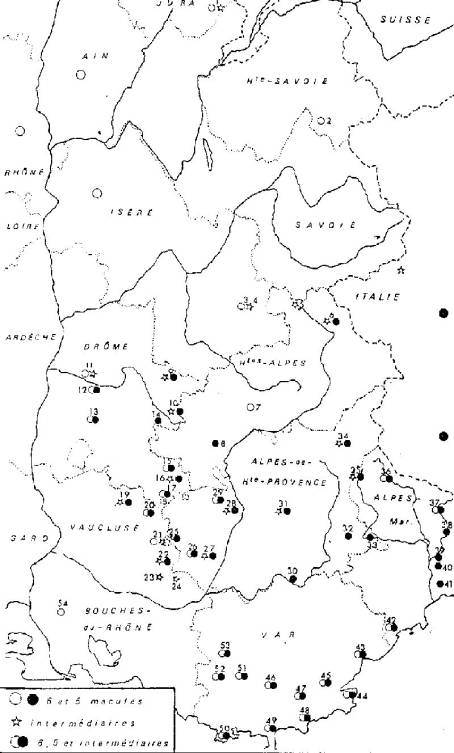
Répartition
qualitative des formes de Z. filipendulae L.
dans le sud-est de la France.
Jura : 1,
Lamoura. - Haute-Savoie : 2, Megève. - Isère : 3, Le Bourg-d'Oisans; 4, Les
Deux-Alpes. - Hautes-Alpes : 5, col du Lautaret; 6, montée du Granon; 7, Gap;
8, col de Fay. - Drôme : 9, col de Menée; 10, col de Cabre; 11, Crest; 12,
forêt de Saou; 13, Dieulefit; 14, col du Fay; 15, col de Perty; 16, col
Saint-Jean; 17, Séderon et montagne de Bergies; 18, col de la Croix de l'Homme
Mort. - Vaucluse : 19, Mont Ventoux; 20, Sault; 21, Lagarde; 22,
Saint-Martin-de-Castillon; 23, Grand Luberon; 24, Vitroles. -
Alpes-de-Haute-Provence : 25, Simiane-la-Rotonde; 26,
Saint-Michel-l'Observatoire et col de Val-Martine; 27, Dauphin; 28, montagne de
Lure; 29, forêt de Valbelle; 30, col d'Ayens; 31, Digne; 32, Annot; 33,
Entrevaux; 34, Barcelonnette. - Alpes-Maritimes : 35, Esteng; 36, Auron; 37,
Valdeblore, La Colmiane, Saint-Martin-Vésubie, Venanson; 38, Lantosque; 39,
Lévens; 40, Saint-Blaise, mont Cima; 41, mont Pacanaglia, Nice; 42, Ranguin,
Mandelieu. - Var : 43, Fréjus, Villepey, Roquebrune; 44, Saint-Tropez; 45, La
Garde; 46, La Foux; 47, Collobrières; 48, Bormes; 49, Hyères; 50, forêt de
Janas, Six-Fours; 51, Mazaugues; 52, Les Glacières, Sainte-Baume; 53,
Saint-Maximin, Seillons. - Bouches-du-Rhône : 54, Saint-Martin-de-Crau.
B - Des
Pyrénées méditerranéennes au sud du Massif Central.
|
Secteurs et localités |
5 taches |
Intermédiaires |
6 taches |
|
Sud-est
des Pyrénées-Orientales Coustouges
(sur plus de 5 ans) Sainte-Colombe,
Camélas Centre
des Pyrénées-Orientales Betllans,
Tarrérach Nord
des Pyrénées-Orientales - Aude Col
Saint-Louis, Col de la Fage, Col de Redoulade, Périllos Larzac,
est de l'Aveyron Le
Coulet, Dargilan, Sainte-Rome de-Dolan, Larvernhe
(près Séverac) Hérault
(près Montpellier) Sète,
Balaruc, Issanka, Cournontéral, Viols-le-Fort Gard Alès
et Bagard |
100% 100% 35 17 0 0 0 |
0 0 12 24 10 5 2 |
0 0 0 23 20 13 15 |
Proportion des formes récoltées dans quelques stations caractéristiques.
Interprétations.
La localisation générale des formes intermédiaires
désigne évidemment celles-ci comme produit mixte de la rencontre des
peuplements à cinq ou six macules.
Le brassage des caractères morphologiques s'observe sur
des dizaines, voire des centaines de kilomètres et aucune restriction de
fertilité ne se manifeste sur le terrain.
En élevage, les formes à cinq ou six macules se sont
révélées pures; en revanche, des individus à six taches, cinq taches un quart
et cinq taches ont été obtenus simultanément de la ponte d'une femelle à cinq
taches et demi. La réalisation de ces combinaisons intermédiaires multiples
désigne a priori un système
polygénique, probablement assez simple, mais une étude expérimentale suivie
demeure indispensable pour définir le mode héréditaire régissant le nombre de
macules.
Si l'expérimentation réalisée ne présente encore qu'un
caractère exploratoire, elle n'en confirme pas moins les observations de
terrain : les formes à cinq ou six macules sont bien cospécifiques.
Au sein des populations naturelles, le brassage génique
lié à la panmixie semble assurer la circulation des allèles, puisque la
réduction de la sixième macule, par exemple, se retrouve généralement très loin
de tout peuplement à cinq taches. Les cartes dressées pour la zone de contact
des Pyrénées méditerranéennes ou pour le sud-est, indiquent clairement cette
diffusion du caractères "cinq taches" à partir de l'Espagne ou de
l'Italie dans le peuplement à six taches du reste de l'Europe.
La persistance de quelques populations à six macules,
essentiellement localisées sur le littoral, semble confirmer cette
interprétation. Tels apparaissent les peuplements de Sète,
Villeneuve-lès-Maguelone, puis de Fréjus, des étangs de Villepey, Saint-Tropez,
Hyères, etc. (sous-espèce callimorpha
Dujardin). Toutes ces stations, liées aux zones humides littorales, se trouvent
sub-insularisées, au même titre que le lido de Venise, où vole également une
grande forme à six taches. Toutes sont aussi menacées de destruction par
l'urbanisme...
ISOLEMENT ET DIFFÉRENCIATION DES POPULATIONS
1 - L'adaptation divergente
chez les Melanargia ou
"Demi-deuil". (Fiche 5)
Interprétation :
Les populations de M.
galathea les plus claires existent en Normandie, les plus sombres dans le
sud-est de la France, l'Italie et la Grèce : la mélanisation de l'espèce semble
suivre le gradient des températures.
En revanche, M.
lachesis confiné à la péninsule ibérique et au Languedoc-Roussillon
apparaît toujours plus clair que M.
galathea. Il se serait adapté aux biotopes xériques et chauds par
albinisme, la réduction des macules noires entraînant une moindre absorption de
chaleur.
Remarque : Chez M. galathea les écailles mélanisées réduiraient la perte d'eau (?).
Cependant dans le midi de la France, cette espèce fréquente plutôt les endroits
relativement frais, les bois clairs, les lisières et allées forestières, etc.
En revanche, M. lachesis vole au
plein soleil estival.
Dans l'échantillon du col de Grès, les hybrides ne
représentent guère plus du 1/5 du total et le contact entre les deux Melanargia n'excède pas quelques km.,
situation très différente de celle observée pour les formes de Z. filipendulae. De plus aucun caractère
de l'un des deux taxons ne se retrouve chez l'autre. (Pas d'introgression). La
faiblesse des échanges géniques démontre qu'il s'agit donc de deux espèces bien
qu'elles ne soient pas totalement séparées par une interstérilité stricte. (Pas
de barrière pré-copulatoire). Ce résultat semble logique puisque l'hybride
réalise une combinaison désavantageuse par rapport à l'un ou l'autre type
adaptatif des deux espèces. La spéciation résulterait donc ici d'une divergence
adaptative.
2 - Les adaptations
trophiques chez les "Damiers". (Fiche
6)
Interprétation :
Considérée globalement, l'espèce E. aurinia se révèle plus ou moins polyphage, au moins oligophage.
La monophagie des différentes populations pourrait être alors comprise comme
exploitation de la plante-hôte possible présente dans tel ou tel biotope. Les
résultats des élevages des hybrides infirment ce raisonnement; ils démontrent
que la spécificité trophique procède d'un déterminisme génétique.
Il est raisonnable d'admettre qu'E. aurinia a conquis ses divers biotopes de proche en proche, en
particulier en altitude, en colonisant dans chacun la plante-hôte compatible
avec ses exigences trophiques et ses possibilités métaboliques. Le végétal a du
servir de filtre écologique, sélectionnant les individus aptes à se développer
à la fois sur la nouvelle source de nourriture et dans un biotope nouveau. Les
pressions de sélection exercées par les différents milieux ont ensuite conduit
à la réalisation des diverses sous-espèces. Celles-ci demeurent cependant
interfertiles et le palier spécifique n'est donc pas atteint.
Les espèce voisines, des genres Euphydryas et Melitaea,
demeurant tributaires des mêmes contraintes trophiques, ont du suivre le même
processus de diversification adaptative mais sont en outre parvenues à
l'isolement reproductif. Le problème majeur reste alors d'établir si cet
isolement résulte d'une évolution quantitative par l'accumulation de mutations
sélectionnées par le milieu et conduisant à l'incompatibilité génétique ou si
celle-ci se trouve acquise à un moment quelconque, qualitativement, par
modification d'un ou quelques gènes cruciaux. Dans cette dernière éventualité,
évolution adaptative et spéciation peuvent être totalement indépendantes.
Documents complémentaires.
La réalité des affinités trophiques pour les genres Euphydryas et Melitaea peut être aisément établie en rapportant la liste des
plantes-hôtes de ces papillons à la classification botanique de L. Emberger ou
d'autres systèmes phylétiques actuels. (p.
et ).
Remarques complémentaires :
Les divergences adaptatives semblent évidentes lorsque
l'on rapproche les sous-espèces d'E. aurinia entre elles. Ainsi E. aurinia beckeri Led. est une grande
forme brillamment colorée dans les tons rouges, répandue en Espagne et dans le
Roussillon en France; elle vit à faible altitude sur quelques Chèvrefeuilles (Lonicera etrusca et L. implexa en particulier). E.
aurinia pyrenes-debilis Verity,
petit, très sombre, fréquente les pelouses subalpines de l'est pyrénéen, vers
2000-2500 m., et ses chenilles vivent surtout sur Gentiana alpina. Ces deux sous-espèces produisent des hybrides expérimentaux
fertiles et abondants au-delà de la F1. Un hybride très voisin, E. aurinia kricheldorffi Collier existe
d'ailleurs naturellement dans les Monts Cantabriques. Les pressions de
sélection dissemblables, très fortes aussi bien en altitude qu'en zone
méditerranéenne, n'ont donc pas altéré l'interfertilité. Enfin, d'autres
considérations laissent penser que ces deux taxons sont restés longtemps
isolés...
Si la séparation des Demi-Deuil peut résulter d'une
divergence adaptative, les aurinia
montrent que la réciproque ne se vérifie pas : la diversification induite par
l'environnement n'implique pas nécessairement l'incompatibilité génique...
L'exemple suivant, fondé sur un autre mode de diversification, complétera ces
observations.
3 - Variations des pièces
génitales chez la Noctuelle de l'Aubépine, Allophyes
oxyacanthae L.
(Fiche 7)
Interprétation :
L'aspect extérieur du papillon assure la protection
contre les prédateurs; coloration et graphisme cryptiques sont donc
vigoureusement sélectionnés partout où existe l'espèce. En revanche, les valves
ne sont pas soumises à la même pression de sélection et toute mutation peut
être indifféremment conservée si elle n'entrave pas les facultés procréatrices.
Des espèces morphologiquement semblables peuvent ainsi différer
considérablement par leurs "genitalia" (ensemble des pièces
copulatrices).
La configuration de la valve droite de corsica apparaît sensiblement
intermédiaire entre oxyacanthae et alfaroi. Or cette structure se localise
précisément dans la zone du contact géographique entre ces deux derniers
taxons. De plus la recombinaison des caractères traduit un échange génique sans
restriction entre les différentes populations en contact. A. oxyacanthae et A. alfaroi
ne sont donc pas isolés génétiquement et les formes de type corsica ne constituent pas une entité
distincte. Cependant la rupture des flux géniques, du fait de l'insularité de
la Corse, a pu conduire à la réalisation d'un type plus homogène dans cette
île.
Conclusions.
La sélection adaptative s'exerce indépendamment et à des
vitesses variables sur les caractères d'une espèce; ceux soumis à des pressions
de sélection fortes et constantes varient très peu... Ces mécanismes,
concomitants de la spéciation, peuvent interférer avec cette dernière mais sans
lien nécessaire. La séparation spécifique n'intervient que si l'interfertilité
n'est plus possible ou conduit à des résultats désavantageux.
PLANTES NOURRICIÈRES DES EUPHYDRYAS ET DES MELITAEA
|
Plantes-hôtes des chenilles |
Principales espèces ou sous-espèces Europe Amérique |
|
|
CAPRIFOLIACÉES - Chévrefeuilles - Symphorine DIPSACACÉES - Succise - Scabieuses - Knautie - Cephalaire GENTIANACÉES - Gentiane alpine - Gentiane de Koch - Gentianes diverses BORRAGINACÉES - Mertensis
ciliata SCOPHULARIACÉES - Molène - Linaires - Linaires,
Véroniques, Mélampyre - Scrophulaires
américaines diverses - Castilleja, Orthocarpus Collinsia,
Pedicularis - Chelone
glabra - Pentstemon - Besseya
alpina PLANTAGINACÉES - Plantains divers - Plantain alpin |
E. intermedia E. aurinia beckeri E. aurinia
aurinia E. aurinia
pyrenes-debilis (en partie < 2000 m) E. aurinia
xeraurinia E. aurinia
xeraurinia E. aurinia
provincialis E. desfontainii E. aurinia
pyrenes-debilis E. aurinia
pyrenes-debilis E. aurinia glaciegenita M. varia M. trivia M. deione M. didyma; M. diamina M. aurelia;
M. deione; etc... M. cinxia; M. didyma; M. diamina; M. athalia; M.parthenoides;M.
aurelia; M.
britomartis M. cynthia |
E. chalcedona E. chalcedona
sierra E. chalcedona Différentes populations d'E. editha E. phaeton E. anicia, E.
colon E. anicia en
altitude Différentes populations d'E. editha
et E. phaeton |
Note : Les Dipsacacées n'existent pas en Amérique (Famille
eurasiatique).
(Les principales familles n'ont été notées que dans la lignée IV)
Cet auteur a proposé une classification des végétaux
supérieurs en plusieurs lignées indépendantes établies à partir de
l'organisation comparée des parentés chimiques, des données paléontologiques,
de la biogéographie, etc...
ASPECTS GÉNÉTIQUES DE LA
SPÉCIATION
1 - Structuration du génome.
(Fiche 8)
Interprétation :
Le dénombrement des chromosomes de certains caryotypes de
Polyommatus évoque une polyploïdie :
24 - 45 - 88. Des remaniements chromosomiques par fusion, fractionnement,
transfert, etc., doivent intervenir mais le type holocentrique des chromosomes
ne facilite pas l'étude de ces phénomènes...
La pulvérisation du génome augmente les possibilités de
recombinaison de manière exponentielle et confère une plus grande liberté aux
différents gènes. Ces facteurs favorisent la diversification des peuplements et
ouvrent un large champ à la sélection adaptative, peut-être à des
restructurations stables du génome (?). L'ensemble de ce groupe paraît
actuellement en pleine crise de spéciation.
Les plaques métaphasiques des hybrides présentent des
appariements variables, ce que traduisent les numérations chromosomiques et les
formes en croissant, très atypiques, visibles sur les figures. Cependant la
production d'hybrides féconds après une ou deux générations entre espèces à
formules chromosomiques très différentes montre que l'information génétique
conserve une certaine cohérence en dépit des morcellements et réarrangements
chromosomiques.
2 - Mutations.
Mutation rousse chez Euphydryas aurinia. (Fiche 9)
Interprétation:
Le caractère réapparaissant à chaque croisement même avec
des souches très différentes de celle d'origine, est nécessairement dominant.
Tous les individus roux hybridés possédaient un génotype hétérozygote Rn, les
autres une combinaison nn, comme le montrent les proportions des formes
normales et rousses, d'environ 50%. Enfin, le gène muté n'apparaît pas lié aux
gonosomes puisque transmis indifféremment par un mâle ou une femelle dans les
mêmes proportions.
Une dépression numérique par consanguinité est évidente
dans les émergences de 1987.
De manière générale, les résultats observés obéissent aux
lois du monohybridisme mendéléien : la mutation pourrait concerner un gène
contrôlant la fin de la chromatogenèse...
La modification produite, très accusée, s'apparente à une
"grosse mutation", rarement observée en milieu naturel et presque
jamais persistante. Ce type de mutation n'a pratiquement pas d'incidence dans
la spéciation.
Documentation complémentaire
: Mutation "sans diapause".
Le cycle biologique d'Euphydryas
aurinia, à faible altitude, s'établit typiquement comme suit :
- Les imagos émergent en mai.
- La ponte éclôt après un mois d'incubation environ;
- les jeunes chenilles tissent un nid de soie et ont une
activité grégaire jusqu'en juillet-
août;
- elles entrent alors en diapause et ne reprennent leur
activité qu'en février-mars, en se dispersant à leur dernière mue.
- Chrysalide en avril.
En août 1978, 10 chenilles d'un élevage provenant de
Lozère ne sont pas entrées en diapause et ont continué à se nourrir. Elles ont
produit des chrysalides puis des imagos en octobre et quatre pontes ont été
obtenues, écloses en novembre 1978. L'élevage a été poursuivi en laboratoire et
une deuxième génération de papillons (31 mâles et 37 femelles) a émergé courant
janvier 1979, fournissant une ponte partiellement fertile. Les jeunes larves se
sont dispersées et sont mortes début mars 1979 en conséquence probable de la
consanguinité et de la nature inadéquate de la nourriture utilisée en hiver...
L'observation établit ainsi la réalité d'une mutation
modifiant une caractéristique biologique de l'espèce, ici la diapause. Le
passage d'une race univoltine à une forme plurivoltine semble donc aisément
réalisable et constitue une potentialité adaptative évidente.
3- Polymorphisme intra et
inter-spécifique.
Le polymorphisme
phénotypique au sein de l'espèce :
l'Argus bleu-nacré (Polyommatus coridon, Poda). (Fiche 10)
Interprétation :
Aucune corrélation simple avec des facteurs climatiques
ou géographiques ne semble expliquer la répartition des femelles bleues. La
valeur adaptative de la mutation paraît nulle.
Même si certains caractères d'une population paraissent
neutres dans les circonstances spatio-temporelles où ils sont observés, ils
constituent une réserve dans laquelle l'espèce pourra puiser si elle doit faire
face à un changement de son environnement. Le polymorphisme permet encore la
conquête de nouveaux biotopes par les individus porteurs des caractères les
mieux adaptés au sein de l'espèce.
Remarque :
Le phénotype "femelle
bleue" étant récessif doit se trouver vigoureusement sélectionné pour
atteindre des proportions importantes, voire 100%. Il pourrait être lié à un
autre gène conférant un avantage sélectif ou dépendre de comportements propres
à l'espèce : les mâles pourraient "préférer" les femelles bleues,
hypothèse à tester expérimentalement.
Cependant la pigmentation alaire chez les Lépidoptères
est tributaire d'un petit nombre de gènes, parfois un seul, il est donc
possible que la mutation bleue ne soit pas apparue dans le peuplement du
sud-est de la France, d'origine biogéographique différente de celle de
l'ouest...
Le polymorphisme génotypique
intraspécifique. (Fiche 11)
Interprétation :
Les histogrammes traduisent la diversité génotypique des
populations d'une même espèce, soit qualitative : elles ne renferment pas
toutes les mêmes allèles et rarement leur totalité; soit quantitative : les
proportions diffèrent pour les allèles qu'elles possèdent en commun, certains
demeurant rares...
De telles analyses renseignent sur les parentés
phylétiques de l'ensemble du peuplement et sur l'importance des échanges
alléliques entre populations géographiquement proches. Les fortes disparités
indiquent que la divergence par mutation et dérive des petites populations se
traduit rapidement dès que s'instaure une entrave aux flux géniques
interpopulationnels. Un échantillonnage plus serré permettrait l'approche des
comportements et de l'écologie propres à chaque espèce...
La diversification génotypique des populations étend les
potentialités adaptatives et évolutives d'une espèce par opposition à la
relative homogénéité réalisée par la panmixie du peuplement, c'est à dire le
brassage génétique entretenu sans restriction entre tous les individus.
L'espèce risque cependant l'émiettement, la perte de son identité et
l'extinction des populations qui ne possédent pas une réserve allélique
suffisante en cas de modifications de leur environnement.
Le polymorphisme génotypique
interspécifique. (Fiche 12)
Interprétation :
Pour chaque gène, les trois espèces se séparent nettement
soit par leurs proportions alléliques (PGM), soit par la possession d'allèles
différents ou caractéristiques (MDH). En moyenne, L. bellargus révèle des loci moins polymorphes que chez les deux
autres espèces.
Il est clair que les trois espèces ne disposent pas des
mêmes potentialités adaptatives et évolutives.
Les comparaisons permettent ici la séparation des espèces
surtout lorsqu'un locus diagnostic apparaît, comme l'allèle c' de la MDH chez L. bellargus. L'intérêt est
particulièrement fort pour les populations sympatrides d'espèces très proches
qui tendent à réaliser des phénotypes adaptatifs convergents.
La variation géographique des fréquences alléliques peut
encore traduire la réalisation d'échanges géniques interspécifiques exprimant
alors le degré de parenté des espèces, c'est à dire l'ancienneté de leur
séparation. Dans l'exemple retenu, L.
bellargus paraît plus éloigné des deux autres espèces très proches l'une de
l'autre.
4 - Une stase évolutive dans
le genre Euphydryas. (Fiche 13)
Interprétation :
L'aire totalement disjointe d'E. iduna et la dispersion extrème, de l'Europe à l'Amérique du
Nord, d'autres espèces du même genre résultent certainement d'épisodes
multiples d'extension, de régression et de migration. Par voie de conséquence,
et compte tenu des distances géographiques considérables, il est logique
d'admettre que ces populations sont séparées de longue date du peuplement
ancestral. La quasi-identité de forme des iduna
du Caucase, de la péninsule scandinave ou de Mongolie traduit alors une fixité
surprenante des caractères de l'espèce qui semble ne pas répondre aux pressions
environnementales qu'elle subit. On nomme stase cet arrêt évolutif.
De telles observations semblent traduire une certaine
homéostasie génétique, une forte cohérence de l'ensemble du génome : les gènes
étroitement coaptés constitueraient une entité fonctionnelle éliminant les
variations moins performantes. Il demeure possible cependant que la variation
mutationnelle existe comme ailleurs mais qu'elle ne touche pas de caractères
aisément perceptibles...
Il est intéressant de rapprocher l'apparente fixité de
ces peuplements disjoints de l'uniformité présentée par les espèces
migratrices, quasi-identiques sur l'ensemble de la planète pour certaines,
alors que l'on regarde généralement dans ce cas le brassage panmictique
qu'elles réalisent comme cause d'homogénéité. Les deux processus ne se contredisent
cependant pas et peuvent conduire indépendamment à des résultats analogues.
SPÉCIATION COMPLEXE DANS LE
GENRE EREBIA
Les connaissances acquises dans ce genre ont permis de
séparer progressivement les espèces, parfois très proches comme elles le sont
dans le groupe d'E. tyndarus. Les
deux fiches (non numérotées) consacrées à ce complexe spécifique réunissent
l'essentiel des données qui permettent d'établir le scénario général de la
spéciation probable du groupe.
Les éléments essentiels à considérer dans ce processus sont
repris ci-dessous.
·
La
mélanisation poussée étant commune à toutes les espèces du genre Erebia, l'adaptation aux fortes
altitudes est certainement antérieure et indépendante de la spéciation étudiée.
·
Les
similitudes morphologiques, y compris celle des valves de l'appareil copulateur
des mâles, suggèrent une parenté proche entre les taxons considérés, non de
simples convergences adaptatives.
·
Les
disjonctions d'aires observées actuellement doivent résulter de plusieurs
cycles d'extension-régression qui ont permis l'isolement des populations puis,
après différenciation, leur migration pendant les périodes de refroidissement.
Le parallèle exact avec les fluctuations climatiques reste à établir...
·
Cohabitation
et interstérilité montrent que le palier spécifique doit être atteint pour tous
les taxons. Cependant ceux-ci occupent les mêmes niches écologiques, à une
altitude donnée, et la compétition interspécifique limite ainsi la sympatrie.
·
La
forme des valves traduit l'aspect aléatoire des mutations et constitue un
caractère de faible valeur adaptative, neutre vis-à-vis du milieu et sans
incidence tant que l'accouplement n'est ni entravé ni favorisé de manière
significative. Il en va probablement de même pour les faibles variations
relevées dans l'ornementation alaire.
·
Les
remaniements chromosomiques, par fusion ou morcellement, jouent certainement un
rôle fondamental dans l'isolement reproductif. Les valeurs n=25 et n=51 pour hispania et iranica évoquent une diploïdie possible que semble cependant
contredire l'éloignement géographique...
·
Les
populations d'E. cassioides, les plus
répandues et les plus dispersées géographiquement, peuvent traduire la
disjonction la plus récente et demeurent les meilleures candidates à la poursuite
de cette évolution.
Les analyses
électrophorètiques récentes confirment les fractionnements successifs subis par
le peuplement d’E. cassioides et la
réalité de la divergence allélique des isolats. Elles établissent encore l’intérêt
des études biogéographiques fines permettant de comprendre l’histoire des
peuplements. Cependant les coupures repérées n’apportent aucune indication
directe sur l’interfertilité ou l’interstérilité des populations et donc sur le
palier de spéciation atteint. La fixation taxonomique au niveau subspécifique
ou spécifique demeure conventionnelle pour les ensembles allopatriques et
fondée ici sur la comparaison des distances génétiques aux taxons voisins. Tel
est encore le problème posé par E.
hispania de Sierra Nevada dont les alléles se séparent nettement de ceux du
peuplement pyrénéen, indiquant une rupture ancienne des échanges géniques entre
ces deux entités.
RÉFLEXION SUR LA NOTION D'ESPÈCE ET SUR LA SPÉCIATION
LE CONCEPT DE L'ESPÈCE.
Les publications consacrées à ce sujet constituent une
masse énorme de documents et d'opinions que nous n'avons ni la possibilité, ni
la prétention, de présenter ici. Un survol de l'évolution de cette notion se
révèle particulièrement instructif.
La désignation binominale due à Linné, traduit le concept
typologique de l'espèce : tout ce qui est conforme à l'exemplaire-type pris en
référence appartient à l'espèce. Cette conception demeure à l'origine de la
référence muséologique qui permet d'établir la nomenclature en accord avec les
principes d'antériorité et autres règles nomenclaturales. Il s'agit du
consensus international minimal, d'ordre administratif en quelque sorte,
servant de garantie et de garde-fou... L’identification par l’image procéde
encore du même esprit.
Le critère de fécondité a été introduit avec Buffon et
Cuvier : l'espèce est la collection des corps organisés, nés les uns des autres
ou de parents communs, et de ceux qui leur ressemblent autant qu'ils se
ressemblent entre eux. Pendant longtemps, l'accroissement des connaissances n'a
fait que diversifier les critères structuraux, morphoanatomiques et
mixiologiques de l'espèce sans remise en question fondamentale.
Ecologie et génétique des populations ont transféré ces
concepts au niveau des populations, transformant ainsi la notion abstraite
d'espèce en un ensemble territorialement limité constitué par un pool de gènes
isolé de ses voisins.
Enfin les ponctualistes, se fondant sur la paléontologie,
ont mis en exergue la durée de l'espèce dans son cadre géologique; l'espèce est
une stase dans un équilibre intermittent...
Nous retiendrons donc les notions paraissant
fondamentales et classiques : l'espèce peut être définie à partir des critères
de similitude morpho-anatomique, d'interfécondité et d'identité génétique. Pour
certains cependant, l'espèce demeure indéfinissable et seuls les individus qui
la composent sont capables de se reconnaître entre eux ! Malgré ce, seule
l'espèce est généralement considérée comme unité naturelle fondamentale par
opposition aux taxons infraspécifiques et surtout supraspécifiques, tels le
genre et le sous-genre, purement conventionnels.
En fait, la variation est continue : il n'y a pas de
différence de nature entre forme individuelle, morphe, sous-espèce,
quasispecies ou semispecies et la multitude des catégories infraspécifiques qui
ont été distinguées. Selon les fonctions qu'elles affectent, les mutations
n'ont pas le même impact sur l'organisme et la sélection, influencée par une
infinité de facteurs, diversifie à l'extrême les conséquences qu'elles
engendrent. Tous les intermédiaires existent dans les degrés de séparation,
jusqu'au niveau supraspécifique, et défient les tendances à la rationalisation
de l'esprit humain, tenant en échec l'impérieux besoin de classement que fait
naître chez l'entomologiste l'objet même de son étude...
LE MODÈLE ALLOPATRIQUE DE LA
SPÉCIATION NÉODARWINIENNE.
L'utilisation du terme spéciation ne se développe, en
Lépidoptérologie en particulier, qu'avec la seconde moitié du XXème siècle.
Elle traduit une meilleure perception de la disjonction d'une espèce initiale
en deux nouvelles espèces, phénomène jusque-là inclus dans les processus
évolutifs généraux, assez flous. Le modèle proposé intervient ici en conclusion
de l'étude des quelques situations analysées précédemment.
La persistance d'une espèce est assurée par le flux
génique qu'entretient la reproduction des individus qui la composent, que
ceux-ci constituent une population unique ou un peuplement plus ou moins
dispersé. Toutes modifications géographiques, climatiques, comportementales,
etc, supprimant les contacts entre populations créent des isolats qui évoluent
alors séparément.
De nombreux mécanismes, souvent recombinés de manière
complexe, peuvent rendre compte de la divergence évolutive observée : le
polymorphisme du peuplement initial engendre une répartition inégale et
aléatoire des allèles dans les isolats (effet fondateur) qui s'accompagne d'une
dérive génétique mutationnelle d'autant plus importante que la fraction isolée
compte moins d'individus (évolution neutraliste). Les pressions de sélection
exercées par les différents biotopes engagent certaines évolutions dans des
voies adaptatives variées. De plus la diversité des conditions environnementales
édaphiques, climatiques et biotiques induisent des réponses comportementales
extrêmement variables qui justifient la boutade : "la spéciation demeure
toujours un cas d'espèce".
Le schéma général doit donc être regardé comme cas
médian, de plus grande fréquence, et non comme enchaînement d'étapes obligées;
la polyploïdie peut produire une spéciation immédiate, le mécanisme génétique
créant lui-même l'isolement; inversement, des séparations longues et des
divergences adaptatives n'impliquent pas un isolement reproductif; diverses
stases évolutives suggèrent que la dérive génétique ne soit pas inéluctable,
etc.
Si la multiplicité des cas
particuliers ne peut qu'être brièvement évoquée ici, sa réalité bannit toute
interprétation systématisée, trop simplificatrice et surtout finaliste : le
schéma proposé ne constitue pas une clé permettant la traduction standardisée
des exemples proposés... Il convient donc, conformément à sa valeur d'hypothèse
n'excluant pas d'autres interprétations, de l'utiliser avec précaution et
surtout sans chercher à rationaliser de manière schématique les processus
biologiques.
Le retour en contact de populations géographiquement
séparées, engendré à priori par des modifications inverses de celles qui
avaient conduit à l'isolement, révèle l'interfertilité ou l'interstérilité des
individus, c'est à dire leur aptitude à remettre ou non leur génome en commun.
Si le critère paraît objectif et non ambigu, il demeure en fait difficile à
mettre en oeuvre sur le terrain ou expérimentalement et de nombreux exemples de
fertilité partielle ont été mis en évidence, donnant matière à la description
d'une multitude de catégories intraspécifiques et subspécifiques.
L'interfertilité traduit l'absence de spéciation. Il
reste cependant possible de distinguer des sous-espèces si les deux peuplements
demeurent parapatriques, séparés par une frange d'hybridation dans laquelle se
recombinent les caractères raciaux. Du fait de ce brassage génétique, deux
sous-espèces ne peuvent être sympatriques. Inversement l'interstérilité de
fait, sur le terrain, marque une séparation spécifique traduite par la
cohabitation. Dans le cas des espèces jumelles, aucune différence morphologique
directement perceptible n'indique l'appartenance à l'une ou l'autre espèce.
Parmi les nombreuses situations traduisant un échange
génique limité entre deux populations, seule la quasispecies = semispecies Mayr
1978 (nec semispecies Mayr 1940) a été retenue à titre indicatif. Une
interfertilité limitée, à un seul sexe par exemple, généralement dissymétrique,
peut permettre à la population B d'acquérir des gènes de A : c'est le mécanisme
d'introgression.